Footnotes
[1] At one point Popper explicitly calls this "Paley's
problem" (p. 345); but elsewhere in this same paper he refers to
"Paley's and Darwin's problem" (p. 342), which is thus consistent
with my designation of 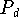 as "Darwin's Problem". I shall
consistently use the latter name; partly because I have used Darwin's
formulation, rather than Paley's, to introduce it; but mainly because
there is a subtle difference between the problem as originally
conceived by Paley, and as actually solved by Darwin. In my
construction ( ), the (conjectured) truth of common descent is an
explicit part of the problem situation; whereas, for Paley, the truth
of common descent was neither a necessary part of (nor even
particularly relevant to) the problem situation.
as "Darwin's Problem". I shall
consistently use the latter name; partly because I have used Darwin's
formulation, rather than Paley's, to introduce it; but mainly because
there is a subtle difference between the problem as originally
conceived by Paley, and as actually solved by Darwin. In my
construction ( ), the (conjectured) truth of common descent is an
explicit part of the problem situation; whereas, for Paley, the truth
of common descent was neither a necessary part of (nor even
particularly relevant to) the problem situation.
[2] To this extent, the title of Darwin's book,
The Origin of Species may be slightly misleading-it directs
attention specifically at the phenomenon of speciation, rather
than at the growth of adaptive complexity. While speciation is
certainly an important issue we can say nonetheless that it is of
secondary importance compared to the question of adaptive complexity.
Diversity of species as such would be of relatively little interest if
all species were equally rudimentary; conversely, even if there were
only one species which displayed adaptive complexity, this one species
would be of overwhelming interest. This is not, of course, to argue
that speciation and the growth of adaptive complexity are
independent phenomena, but
merely that I shall concentrate on the
latter.
[3] Attempts have, of course,
been made to apply "Darwinian" theory to a vast range of domains; I
use the "organismic" qualifier here simply to emphasise that I am
talking about the application of Darwinism in its original context of
the evolution of biological organisms. This usage is loosely related
to the Organismic Evolution of
Beurton (1981), and the Organismic
Selection of Wright (1980).
[4] I find
the latter term to be seriously confusing. This is discussed
at length in (McMullin 1992c).
[5] The common "S" prefixes in
S-lineage and S-value are co-incidental, since they denote
"similarity" and "selection" respectively;
but since only S-lineages can have S-values, it is a convenient
co-incidence; I did not intend it, but I am happy to allow it
stand.
[6] S-value is not even necessarily transitive. One
could formulate S-lineages A, B, C, with properties such that
all three distinct pairs would give rise to selection processes, where
A would reliably displace B, B would reliably displace C, yet
C would reliably displace A!
[7] This thesis is commonly expressed by saying that
variation is "random"; but that term is very vague
and ambiguous.
[8] Admittedly, it is a moot point
whether this is true of unicellular organisms;
but I shall not pursue that further here.
[9] However
note that Popper later repudiated this analysis, as I
discuss in section 5.2.
[10] There may be a lingering misconception that all
tautologies are equivalent, so that the
possibility of "distinct" tautologies cannot arise; this is
not the case. All tautologies have the same truth value
(namely, unconditionally "true"), so that they are
logically equivalent-but this is not at all the same
thing. "Cats are a kind of domesticated feline"
and "  is the ratio of the circumference to the diameter of a circle"
are (qua definitions) two different
tautologies.
[11] See
(Gould 1978, Essay 4) for a more detailed
discussion of whether Darwin "really" endorsed the idea
of necessary progress in evolution.
[12] In this respect, the
term "adaptationist" is unfortunate, as it
invites a form of the tautology misconception 3;
"selectionist" might be less prejudicial.
[13] http://www.eeng.dcu.ie/ tkpw/popconf/popconf.html
[14] http://beast.cc.emory.edu/Jargon30/JARGON_C/CONSHARM.HTML
is the ratio of the circumference to the diameter of a circle"
are (qua definitions) two different
tautologies.
[11] See
(Gould 1978, Essay 4) for a more detailed
discussion of whether Darwin "really" endorsed the idea
of necessary progress in evolution.
[12] In this respect, the
term "adaptationist" is unfortunate, as it
invites a form of the tautology misconception 3;
"selectionist" might be less prejudicial.
[13] http://www.eeng.dcu.ie/ tkpw/popconf/popconf.html
[14] http://beast.cc.emory.edu/Jargon30/JARGON_C/CONSHARM.HTML
Copyright © 1996 All Rights Reserved.
TCR Issue Timestamp: Tue Dec 31 17:37:08 GMT 1996
tcr-editors@www.eeng.dcu.ie